ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ, нуклеотиды, фосфатный остаток которых связан с рибозой в 3'- и 5'-положениях, образуя кольцо; универсальные регуляторы внутриклеточного метаболизма. Открытие циклических нуклеотидов (циклический 3',5'-аденозинмонофосфат, или цАМФ, открыт в 1957 году Э. Сазерлендом и наиболее подробно изучен) — одно из крупнейших научных достижений в области исследования молекулярного механизма действия гормонов. В организме животных цАМФ опосредует действие гормонов, не проникающих внутрь клетки (полипептидных гормонов и катехоламинов), и регулирует множество процессов: синтез и гидролиз гликогена в печени, дифференцировку тканей, кроветворение, тромбоцитоз, явления иммунитета, злокачественного роста, клеточной проницаемости, мышечное сокращение, секрецию гормонов, транскрипцию, трансляцию и многие другое. В стрессовых ситуациях цАМФ служит «сигналом голода». Выделяемый в это время корой надпочечников гормон адреналин при посредничестве цАМФ активирует в клетках печени фермент гликогенфосфорилазу. В кровь выбрасывается большое количество глюкозы и таким образом удовлетворяется острая потребность организма в источнике энергии. Механизм действия цАМФ в клетке связан с активацией цАМФ-зависимых протеинкиназ (ПК) и по существу сводится к фосфорилированию специфических белков (в том числе ферментов), что приводит к изменению их активности и соответствующих функций клетки. Один и тот же гормон в разных тканях вызывает через активацию протеинкиназ фосфорилирование разных белков и обусловливает разные функциональные ответы. У гетеротрофных бактерий цАМФ обеспечивает синтез ряда катаболитных ферментов при отсутствии в среде глюкозы, то есть в условиях голода. Белок, связывающий цАМФ, присоединяется к ДНК и активирует опероны, кодирующие синтез индуцируемых ферментов. У высших растений свет, поглощаемый фитохромом, активирует аденилатциклазу. Из других циклических нуклеотидов известны 3',5'-гуанозинмонофосфат (цГМФ) и 3',5'-цитидинмонофосфат (цЦМФ). Действие цГМФ связано с работой другой независимой системы. Гуанилатциклаза (в основном растворимый фермент), катализирующая образование цГМФ из гуанозинтрифосфата (ГТФ), активируется соединениями, содержащими или образующими свободно-радикальную группу NO (среди них нитроглицерин и другие лекарственные препараты), а также ненасыщенными жирными кислотами, их гидроперекисями, эндопероксидами простагландинов, свободными гидроксильными радикалами. Действие гормонов на активность гуанилатциклазы не связано с взаимодействием их с соответствующими рецепторами, как это происходит в системе цАМФ. цГМФ выполняет самостоятельную, отличающуюся от цАМФ, роль регулятора различных процессов клеточного обмена веществ, однако механизм действия и физиологическая функция цГМФ остаются невыясненными.
Схема механизма действия гормонов у животных с участием цАМФ
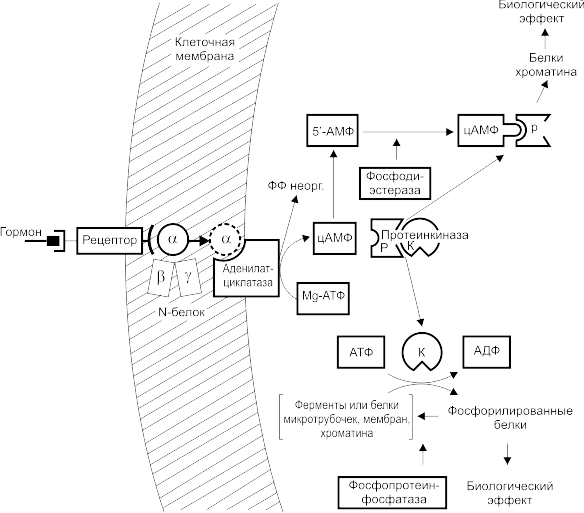
Гормон соединяется с рецептором на внешней стороне клеточной мембраны и при участии N-белка активирует фермент аденилатциклазу (АЦ), локализованную на внутренней стороне мембраны. АЦ катализирует синтез цАМФ из комплекса Mg-АТФ. Образовавшийся цАМФ связывается с ферментом протеинкиназой, в результате чего фермент диссоциирует на регуляторную (Р) и каталитическую (К) субъединицы. К-субъединица фосфорилирует определенные белки, в том числе ферменты, увеличивая или снижая их активность, что приводит к изменению соответствующих функций клетки. Фосфорилирование К-субъединицей белков хроматина или связывание с ним Р-субъединицы приводит к изменению матричной активности хроматина и пролиферативного статуса клетки. На этапе активации аденилатциклазы гормональный сигнал усиливается в 100 —1000 раз, а при активации протеинкиназы ещё в 100 раз, то есть одна молекула гормона может вызвать фосфорилирование 10-5 молекул белка (каскадное усиление). Действие цАМФ в клетке прекращается при гидролизе его фосфодиэстеразой и дефосфорилированин белков фосфопротеннфосфатазой.